Any opportunity to be in the sea is a good opportunity to go jelly-hunting, and the recent participation of HYPNO on a research cruise with the SponGES Project on RV Kristine Bonnevie this late April – early May was no exception!
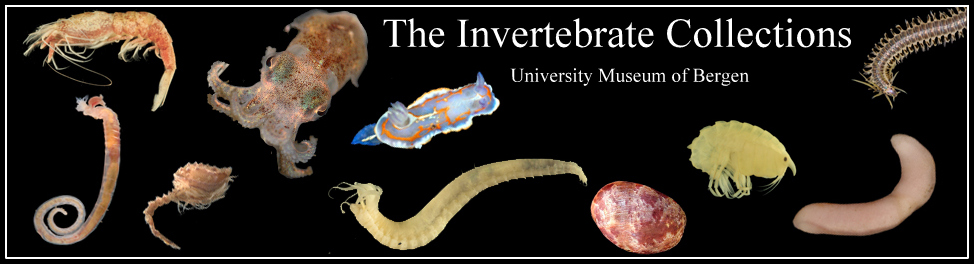
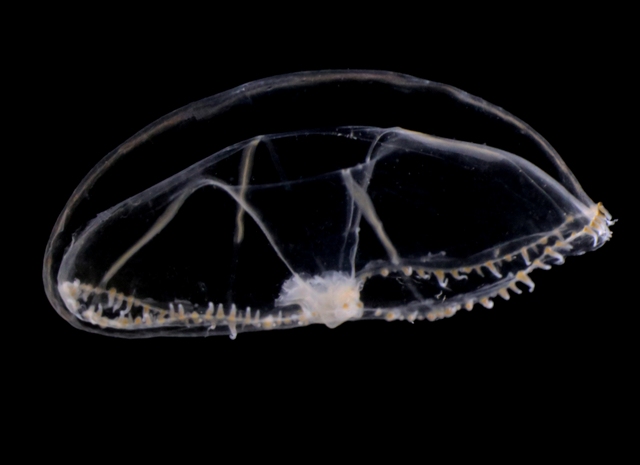
To begin with, we got the chance to sample some hydromedusae and siphonophores with the plankton net in Bømlafjord. As usual, towing the net slowly (~0.3 ms-1) resulted in happy jellies (they get damaged if the net is towed too fast!) that sometimes can be identified with ease. Over 15 different species of pelagic hydrozoans (plus some ctenophores and Tomopteris worms) were present in this vertical tow, with some nice looking critters such as the Eutonina indicans and Leuckartiara octona medusae shown below.
Eutonina indicans
Leuckartiara sp.
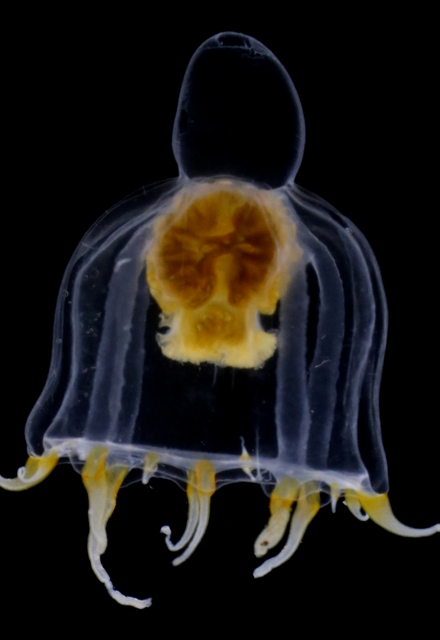
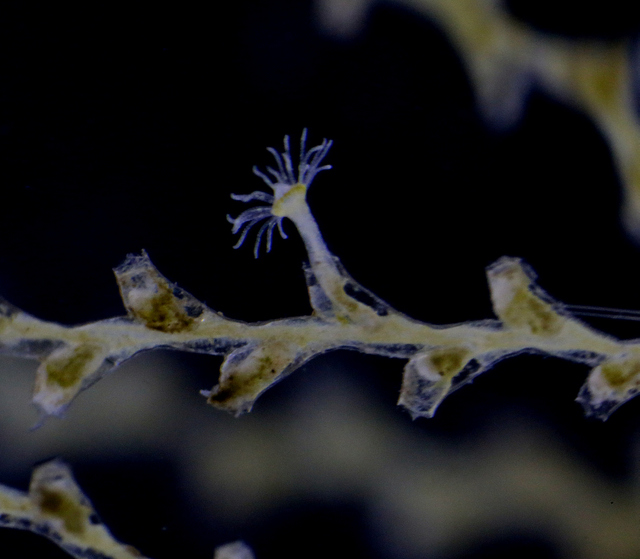
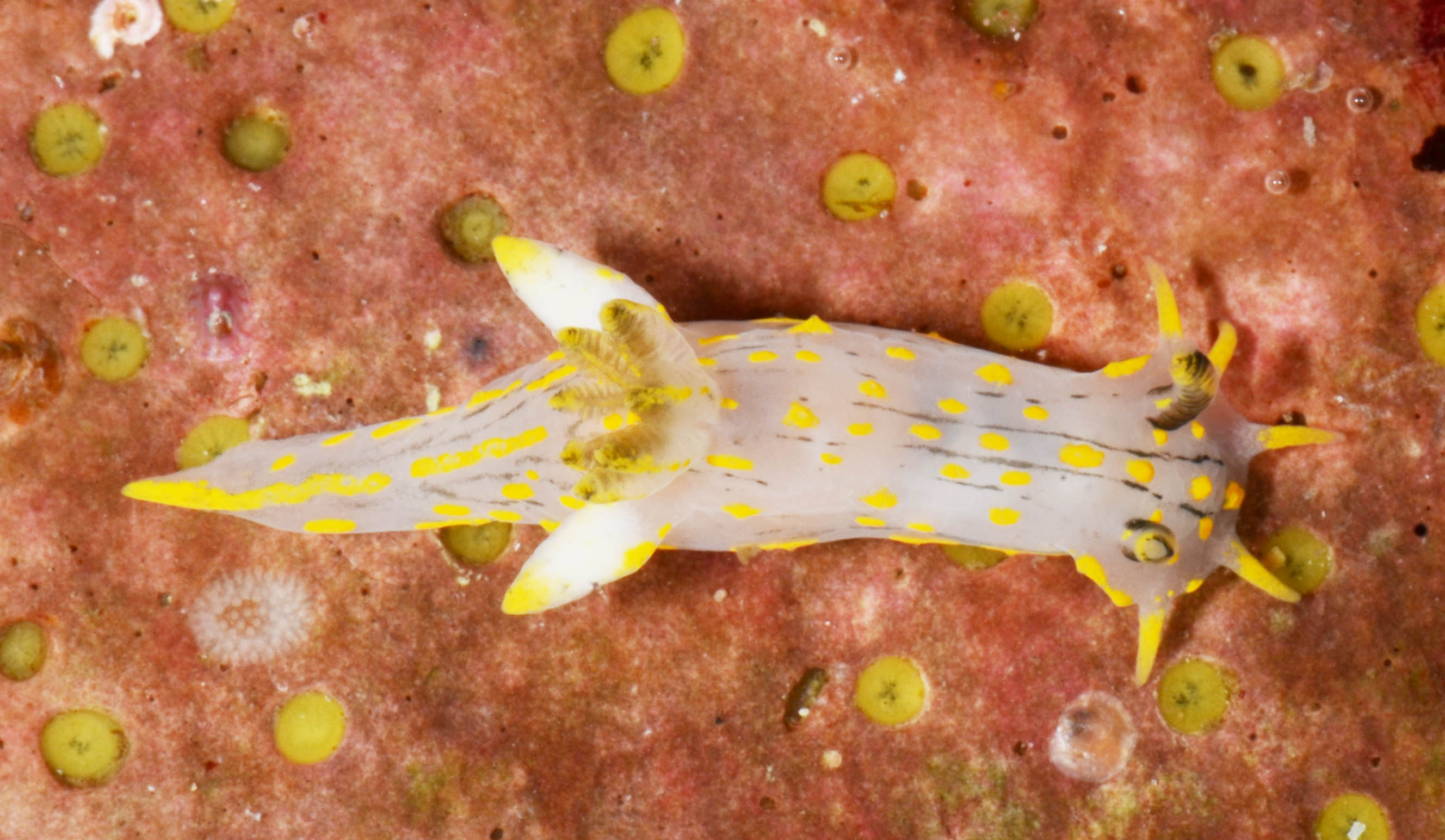
But not only hydromedusae and siphonophores showed up this time; we also got our hands on benthic samples from grabs and trawls, and found hydroids growing on rocks and other sea creatures (mostly sponges and sea squirts). Abietinaria abietina and Sertularella gayi (pictures below) are among the most common hydroids observed so far, and they were hosting a whole bunch of other hydrozoan species growing on top of them: real mini animal forests from the Norwegian waters!
Polycera quadrilineata (Norway) Photo: M. Malaquias
Phillidia ocellata (Mozambique) Photo: M. Malaquias
Chromodoris cf. quadricolor. Vamizi Island
Elysia subornata (Key Largo)
Chromodoris africana (Zavora, Inhambane). This species is part of a complex in need of revision where other “species” imaged here are also part of (e.g. Chromodoris hamiltoni, Hypselodoris regina, Chromodoris elisabethina)
This adventure started 26 years ago, when two Norwegian benthos researchers (Torleiv Brattegard from University of Bergen and Jon-Arne Sneli from the University in Trondheim) teamed up with three Icelandic benthos specialists (Jörundur Svavarsson and Guðmundur V. Helgasson from University of Iceland and Guðmundur Guðmundsson from the Natural History Museum of Iceland) to study the seas surrounding the volcanic home of the Nordic sages. 19 cruises and 13 years later – and not least lots of exciting scientific findings and results the BioICE program was finished.
But science never stops. New methods are developed and old methods are improved – and the samples that were stored in formalin during the BioICE project can not be used easily for any genetic studies. They are, however, very good for examinations of the morphology of the many invertebrate species that were collected, and they are still a source of much interesting science.
Participants of the IceAGE workshop. Photo: Christian Bomholt (www.instagram.com/mcb_pictures)
The dream about samples that could be DNA-barcoded (and possibly examined further with molecular methods) lead to a new project being formed – IceAGE. A large inernational collaboration of scientists organised by researchers from the University of Hamburg (and still including researchers from both the University of Iceland and the University of Bergen) have been on two cruises (2011 and 2013) so far – and there is already lots of material to look at!
Ready to start the workshop! Photo: AH Tandberg
The beaver was here! Photo: AH Tandberg
Ed found the bison! Photo: AH Tandberg
What way should we take? Amphipodologists out of their natural habitat? Photo: AH Tandberg
This week many of the researchers connected with the IceAGE project have gathered in Spała in Poland – at a researchstation in woods that are rumoured to be inhabited by bison and beavers (we didn´t see any, but we have seen the results of the beavers work). Some of us have discussed theories and technical stuff for the papers and reports that are to come from the project, and then there are “the coolest gang” – the amphipodologists. 10 scientists of this special “species” have gathered in two small labs in the field-station, and we have sorted and identified amphipods into the wee hours.
Lauren and Anne-Nina hard at work. Photo: AH Tandberg
Wims microscope after a sample is done.. Photo: Christian Bomholt (www.instagram.com/mcb_pictures)
Lauren after getting the identification right. Photo: AH Tandberg
Ed at work with a nice sample. Photo: AH Tandberg
Lauren examines the specimen while Anne-Nina and Tammy checks the literature. Photo: AH Tandberg
“the Anne-table” in the amphipod lab. Photo: Christian Bomholt (www.instagram.com/mcb_pictures)
It is both fun and educational to work together. Everybody have their special families they like best, and little tricks to identify the difficult taxa, and so there is always somebody to ask when you don´t find out what you are looking at. Between the stories about amphipod-friends and old times we have friendly fights about who can eat the most chocolate, and we build dreams about the perfect amphipodologist holiday. Every now and then somebody will say “come look at this amazing amphipod I have under my scope now!” – we have all been treated to species we have never seen before, but maybe read about. We also have a box of those special amphipods – the “possibly a new species”- tubes. When there is a nice sample to examine, you might hear one of the amphipodologist hum a happy song, and when the sample is all amphipods but no legs or antennae (this can happen to samples stored in ethanol – they become brittle) you might hear frustrated “hrmpfing” before the chocolate is raided.
A large amphipod comes out of the jar! Photo: Christian Bomholt (www.instagram.com/mcb_pictures)
Cleippides quadricuspis. Photo: AH Tandberg
Amphipods sorted and identified. Photo: AH Tandberg
Isopodologists (Martina and Jörundur) visiting the amphipodologists… Photo: AH Tandberg
The samples from IceAGE are all stored in ethanol. This is done to preserve the DNA for molecular studies – studies that can give us new and exciting results to questions we have thought about for a long time, and to questions we maybe didn´t even know we needed asking. We can test if what looks like the same species really is the same species, and we can find out more about the biogeography of the different species and communities.
The geographical area covered by IceAGE borders to the geographical area covered by NorAmph and NorBOL, and it makes great sense to collaborate. This summer we will start with comparing DNA-barcodes of amphipods from the family Eusiridae from IceAGE and NorAmph. They are as good a starting-point as any, and they are beautiful (Eusirus holmii was described in the norwegian blog last summer).
The field-station is ready for easter. Photo: AH Tandberg
The coolest easter-chickens in Spala. Photo: AH Tandberg
Easter-prepared coffee! Photo: AH Tandberg
Happy easter from all the amphiods and amphipodologists!
Anne Helene
Literature:
Brix S (2014) The IceAGE project – a follow up of BIOICE. Polish Polar Research 35, 1-10
Dauvin J−C, Alizier S, Weppe A, Guðmundsson G (2012) Diversity and zoogeography of Ice−
landic deep−sea Ampeliscidae (Crustacea: Amphipoda). Deep Sea Research Part I: 68: 12–23.
Svavarsson J (1994) Rannsóknir á hryggleysingjum botns umhverfis Ísland. Íslendingar og hafiđ.
Vísindafélag Íslendinga, Ráđstefnurit 4: 59–74.
Svavarsson J, Strömberg J−O, Brattegard T (1993) The deep−sea asellote (Isopoda,
Crustacea) fauna of the Northern Seas: species composition, distributional patterns and origin. Journal of Biogeography 20: 537–555.
It certainly does not take a great leap of imagination to get from these Isopoda collected by the MAREANO programme to various science fiction monsters!
click to embiggen!
I just completed photographing and tissue sampling 95 specimens that will be submitted for barcoding through NorBOL – we’ll send them to the CCDB-lab in Canada for sequencing, and upload the metadata and sequences in the BOLD database – fingers crossed for successful sequencing!
Untangling the diversity and evolution of Sea Hares
Aplysia parvula; Føllingen, Norway; Photo by Nils Aukan
Sampling and freezing at Askøy
Dr Carlo M. Cunha from the Metropolitan University of Santos in Brazil (Universidade Metropolitana de Santos), a world expert in the diversity and systematics of Anaspidea heterobranch gastropods, visited the Natural History Museum of Bergen for a month during January/February 2017 to study our scientific collection of these molluscs. The visit was funded by the University of Bergen´s Strategic Programme for International Research and Education (SPIRE).
The Museum holds a large amount of material from the Scandinavian region, but also from the Mediterranean, Macaronesia islands, Caribbean, and western Indian Ocean.
These marine molluscs commonly known by sea hares comprise around 90 currently known species and have long been of major interest to biologists because of their large and easily accessible nervous system, which form the basis of numerous neurophysiological works.
Preserved specimen of Aplysia punctata from Norway
Dissected specimen of Aplysia punctata from Norway
However, the taxonomy of these molluscs and their evolution are still poorly understood. Dr Cunha is using a combination of molecular and morphological tools to learn more about the worldwide diversity of anaspideans and their phylogenetic relationships.
Dr Cunha visit to Bergen has already resulted in the revision and update of the taxonomy of our Anaspidea collection. The Norwegian species of anaspids were revised and redescribed in detail using electron microscopy and DNA barcoding performed in collaboration with Louise Lindblom (University Museum / Biodiversity Labs).
SEM-image of jaws of Phyllaplysia sp from Florida, USA
Additionally several other species from around the world were studied and will be integrated in ongoing taxonomic revisions. Keep tuned!
-Manuel
We’ve also had Lloyd visiting recently, you’ll find a post about that on the Marine Invertebrates of Western Africa blog: click here
A whale recently had to be put down by wildlife management after it had repeatedly beached itself on the island of Sotra outside of Bergen. It was found to be a Cuvier’s beaked whale (Ziphius cavirostris), a species with apparently no official previous records from Norway. The University Museum of Bergen therefore wished to include the whale skeleton in its collections (and future exhibitions, once the remodelling completes).
Arriving at Espegrend
The whale was transported to the Marine Biological Station of Espegrend, and a team of five people from the museum set to work collecting measurements of the whale, taking tissue samples for DNA-barcoding though the NorBOL-project, collecting ectoparasites, and doing photo-documentation.
Collecting measurements
We then began removing the blubber and muscle tissue off the whale so that the bones can be further treated (they contain a lot of oil which needs to be taken care of once the soft tissue has been removed), before the skeleton can be mounted for display.
Starting the work of removing blubber and muscles
Little did we know that what had so far been a local news matter would soon go viral…
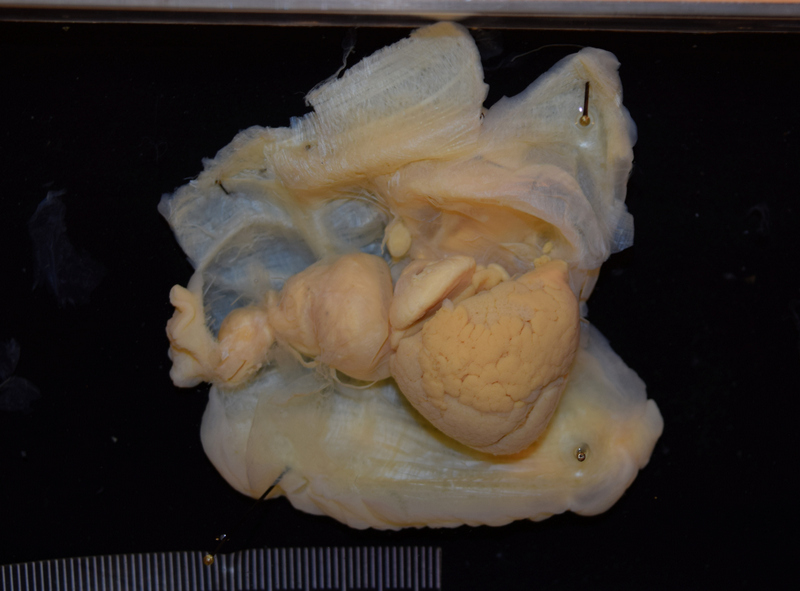
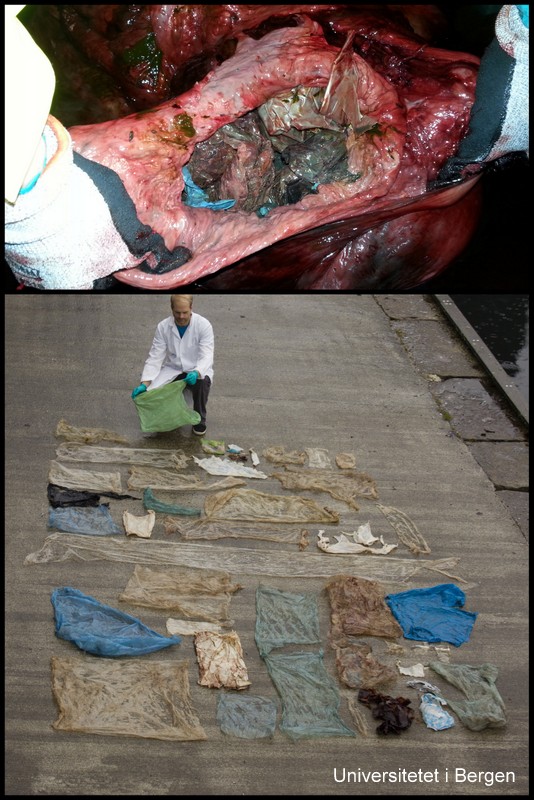
Sadly, it became clear during the autopsy that the whale had been ingesting massive amounts of plastic – as much as 30 plastic bags, and many smaller pieces of plastic. The whale was emaciated, and we believe that the plastic had gathered in such an amount in its stomach that it had created a plug, stopping the digestive process.
The plastic in and from the whale stomach (photos: T. Lislevand, H.Glenner/C.Noever)
The images of all the plastic spread out on the ground became a potent reminder of the tragedies that marine pollution is creating, and has sparked a renewed debate on how we can limit the amount of micro- and macro-plastic that end up in nature.
The news of the whale’s stomach content became international news
What should the Cuvier’s beaked whale have been eating?
Occurring as solitary animals or in small pods, and preferring the deeper open waters, the Cuvier’s beaked whale is not an easy animal to study. We do know that the species have a more or less cosmopolitan distribution, and that it holds the world record for longest and deepest dive for any mammal: one was recorded diving down to 3000 meters.
What data we do have on the species diet comes from beached individuals, and suggests that the species may be a fairly omnivorous predator. From the limited number of Cuvier’s beaked whales that have been examined for stomach content, there are regional differences in the diet, but it seems to consist mainly of cephalopods (squid and octopuses), deep sea fish, and medium sized crustaceans (Santos og andre 2001).
Above are the suckers on the arm of a giant squid, Architeuthis. Below are scars on the skin of a sperm whale. Photo: E.Willassen
The cephalopods appear to be the dominant food source, but this interpretation may be influenced by the longevity of the hard parts of a cephalopod in the stomach.
The tough beaks of a cephalopod consist of chitin, and is used for tearing prey to pieces. Chitin is also found in the suckers of many cephalopods. The beaks can be used to identify the cephalod groups based on their size and shapes. Animals such as jellyfish would be much harder to document as part of the diet, as they would be digested much more rapidly and completely.
We don’t know how well resolved the information produced by the animal’s echo-location is, but it is conceivable that the plastic reflects signals in a way similar to the natural food of the whale, and is therefore “caught” and eaten.
Cephalopod beak, drawing by J.H. Emerton (from Wikimedia commons)
We did find some cephalopod beaks in between the plastic in the whale stomach – so far we have not had the time to attempt to identify these, but we will.
Amongst the plastic there are some cephalopod beaks (dark brown) and a bivalve shell (top left). Photo: C. Noever
The University Museum have extensive cephalopod collections, and long traditions for working with this group – from Dr. Jakob Johan Adolf Appellöf who began working here in 1890, to the material collected in the MAR-ECO project.
MAR-ECO workshop on cephalopoda
From the work of Santos et al 2001 we know that the following species are in the diet of European Curvier’s beaked whales, and are probably amongst the things our whale should have been eating:
Tewuthowenia megalops. Photo: Richard E. Young during MAR-ECO-cruise 2004.
Teuthowenia megalops is an odd squid that floats around in the open water with a propulsion system based on ammoniumchloride that the animal produces by digesting protein. The name “megalops” hints to the huge eyes, which also contain three light producing organs (chromatophores). The species seems to be common in deep water in the north Atlantic (Vecchione et al. 2008). For more information, see Wikipedia.
Mastigoteuthis agassizi
Mastigoteuthis agassizii was originally registered in whale stomachs as Mastigoteuthis schmidti, but from the work on the MAR-ECO project, three species of Mastigoteuthis were considered to all be M. agassizii. Some ambiguity remains about the species of this genus of oceanic squid with a broad distribution in the world’s oceans in depths ranging from 500 to 1000 meters. They have diurnal migration, and may be found hunting closer to the surface at night.
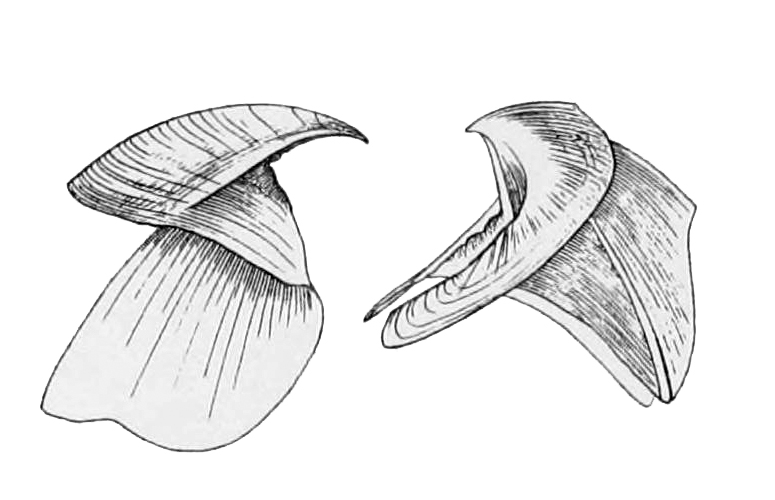
Taonius pavo seen ventrally (above) and dorsally. Illustration from Wikipedia.
Taonius pavo
This little squid is not very well known. It has been recorded from the Atlantic Ocean, but it may have a broader distribution. In this link you will find a video from the Bahamas at 850 m depth where the animal releases bio- luminescent “ink” to confuse a predator and escape.
Histioteuthis bonelli Photographed by Richard E.Young during the Mar-Eco-cruises in 2004
Histioteuthis bonelli, drawing by Ernst Haeckel.
Histioteuthis bonnellii has several names in English, one of which is “umbrella squid”. The name is due to the skirt-like membrane between the arms – when it splays its arms it resembles an umbrella. We don’t know much about the biology of H. bonellii, except that it has several close relatives in the world oceans, and that what has hitherto been considered one species (H. bonellii) may well turn out to be several species.
Todarodes sagittatus
Todarodes sagittatus, the European flying squid, is one of the ten-armed cephalopods that may irregularly occur in schools along the Norwegian coast. T. sagittatus is subject to fisheries.
Vampyroteuthis infernalis
Vampyroteuthis infernalis – the vampire squid is a deep-sea squid with eight arms and a skirt-like mantle between its arms. It also has moveable wings on its body that it can use to manoeuvre with. The name “vampire squid” is not quite true – this is no blood sucker, but it traps organic material from the water masses using long, sticky threads. If threatened, it can invert the “skirt” over its head, resembling a hedgehog. It also has light producing organs towards the back of the body, and can create clouds of bioluminescence. Even with all these defences, it may end up in the stomach of a Cuvier’s beaked whale.
Pelagic crustaceans and deep sea fish are also amongst the recorded prey from Cuvier’s beaked whales. Amongst these we find the fairly large and shrimplike Gnathophausia, found within the order Lophogastrida, which has been studied extensively at the University of Bergen. We also found a bivalve shell in the stomach of our whale, which as far as we are aware of has not been recorded as part of their diet previously.
Plastic or food?
It may seem strange that the whale should ingest large amounts of plastic – why would it do that? If the whale primarily finds its pray by echolocation in the pitch black of the deep sea, it may well be that it is unable to differentiate between the reflected signal from a sheet of plastic, and that from one of its usual prey animals.
Unlike the sperm whales that hunt cephalopods in a similar way, the beaked does not have teeth to grab its pray. Instead they use a suction to ingest the food. Perhaps it is this feeding mode that becomes very unfortunate for the whales in a natural environment with an incredible amount of human garbage.
Hyalinoecia tubicola from the North Sea (by K. Kongshavn).
Quill worms belong to the annelid family Onuphidae and are called like that because of their unique tubes. The tubes are secreted by their inhabitants and are very light and rigid, resembling a quill, the basal part of a bird’s feather used for writing. Quill worms are epibenthic creatures capable of crawling on the surface of the sea floor carrying their tubes along. Their anterior feet are modified, strengthened and enlarged, bearing thick and stout bristles. These anterior feet are used for locomotion.
Quill worms are widely distributed in the ocean inhabiting mostly slope depths down to 2000 m. Being large in body size (up to 10-20 cm long), they can be quite abundant in some areas. Meyer et al. (2016) reported Hyalinoecia artifex reaching up to 70 ind./m2 in the Baltimore Canyon at 400 m water depth. Another quill worm, H. tubicola, which is very common in Norwegian waters, reached up to 272 ind./m2 at 365 m offshore of Chesapeake Bay (Wigley & Emery 1967).
Quill worms are believed to be motile scavengers. Baited monster camera experiments performed at 2000 m deep site in Baja California demonstrated that Hyalinoecia worms can accumulate in hundreds of specimens five hours after the bait (rotten fish) has been deployed (Dayton & Hessler 1972). Myer et al. (2016) analyzed the stable isotope content in Hyalinoecia artifex tissues confirming its secondary consumer status. Their results supported earlier observations on the gut content of the same species by Gaston (1987) showing the presence of the remains of various benthic invertebrates.
Video 1. Quill worm Hyalinoecia tubicola moving inside its tube (by K. Kongshavn).
Video 2. Quill worm Hyalinoecia tubicola protruding from the tube opening. Three antennae and a pair of palps are seen on the head. The first two pairs of feet are enlarged and strengthened (by K. Kongshavn).
Dayton, P.K., Hessler, R.R., 1972. Role of biological disturbance in maintaining diversity in the deep sea. Deep-Sea Research 19: 199–208.
Meyer, K.S., Wagner, J.K.S., Ball, B., Turner, P.J., Young, C.M., Van Dover, C.L. 2016. Hyalinoecia artifex: Field notes on a charismatic and abundant epifaunal polychaete on the US Atlantic continental margin. Invertebrate Biology 135: 211–224. doi:10.1111/ivb.12132
Gaston, G.R. 1987. Benthic polychaeta of the Middle Atlantic Bight: feeding and distribution. Marine Ecology Progress Series 36: 251–262.
Wigley, R.L., Emery, K.O. 1967. Benthic animals, particularly Hyalinoecia (Annelida) and Ophiomusium (Echinodermata), in sea-bottom photographs from the continental slope. In: Deep-Sea Photography. Hersey JB, ed., pp. 235–250. John Hopkins Press, Baltimore.
Last year we had a calendar post about the Heart of the Museum – our type collections.
To recap, a species’ type is “…the objective standard of reference for the application of zoological names. When a new species or subspecies is described, the specimen(s) on which the author based his/her description become the type(s) (Article 72.1). In this way names are linked to type specimens, which can be referred to later if there is doubt over the interpretation of that name.
Consequently types are sometimes referred to as “onomatophores” which means name bearers.”
The location – sampling site – from which the type specimen is described is known as the type locality.
Michael Sars (image from Wikimedia)
As you have probably noticed, polychaetes (bristle worms) are a focus group in our lab, and several species have type localities close by.
The biologist and theologian Michael Sars (1805-1869) lived in the Bergen region for many years. He was a prolific taxonomist, naming 277 species of marine taxa according to the World Register of Marine Species (WoRMS).
Consequently there are quite a few species that have their type locality within easy daytrip-distance by ship for us.
On the hunt with R/V “Hans Brattstrøm”
One such locality is Glesvær, where Michael Sars described several new species in his work of 1835: Beskrivelser og Iagttagelser over nogle mærkelige eller nye i Havet ved den Bergenske Kyst levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes og Molluskernes Classer* (“Descriptions and Observations of some strange or new animals found off the coast of Bergen, belonging to the Classes …”).
The polychaete Amphicteis gunneri (Ampharetidae) is one of these species. It was first described by Michael Sars as Amphitrite gunneri (the species name is an homage to Johan Ernst Gunnerus (1718-1773) who was an active scientist within botany and zoology, as well as the bishop in Trondheim, and one of the founders of Det Kongelige Norske Videnskapers Selskap) in the publication above. Here are his original illustrations of the species:
Amphicteis gunneri by M. Sars (1835)
We have previously submitted several specimens of Amphicteis gunneri for DNA-barcoding through the NorBOL-project – and found that specimens that according to the keys in the literature should all come out nicely as A. gunneri in fact end up in several barcode-based groupings (BINs), meaning that they genetically different from each other. Then we need to unravel which one is the true A. gunneri, and decide what to do with the others. In such cases, material from type localities is invaluable. By sending in specimens identified by resident taxonomists as A. gunneri from the type locality, we hope to figure out which BIN represent A. gunneri, and which represent potentially new species.
We were also able to photograph live specimens showing the nice coloration of this worm. Fixed specimens lose this colour and become uniformly yellow/white (no dots).
Amphicteis gunneri collected at type locality. Photo: K.Kongshavn
Polina Borisova, a first year master student from the Zoological Department of the Moscow State University (Russia), is coming to the Invertebrate Collections of the University Museum of Bergen with a 1-month research visit in January 2017.
Polina is going to work on the bristle worms from the family Lumbrineridae studying the collection from West Africa and Norway. Her project is jointly supervised by Dr. Nataliya Budaeva from the University Museum of Bergen and Dr. Alexander Tzetlin from the Moscow University.
Various Lumbrineridae from West Africa, scale 1 mm (Photos from BOLD).
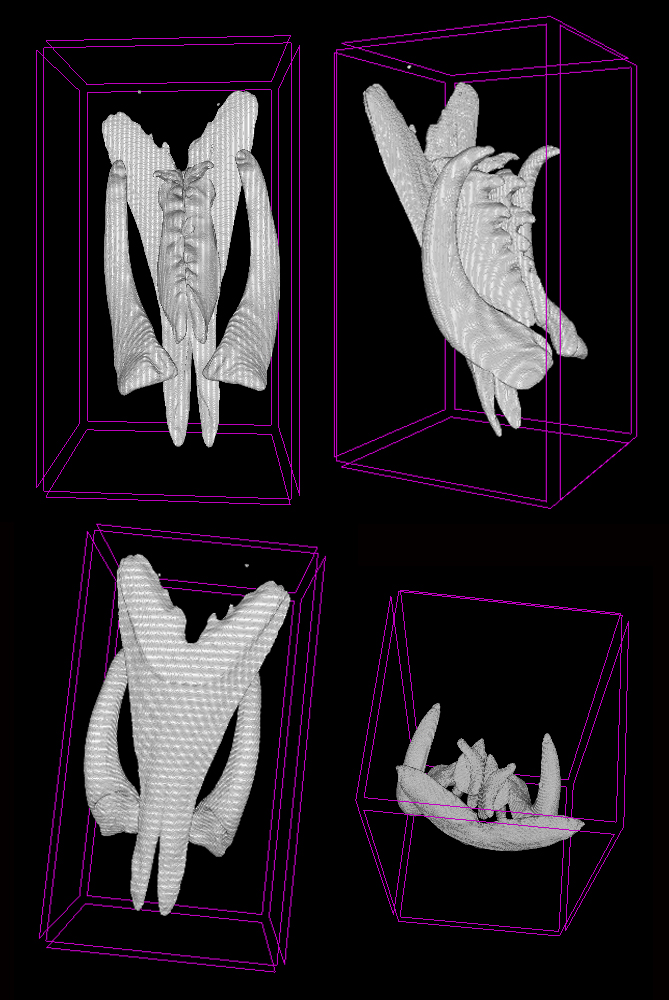
Lumbrineridae are the worms with relatively poor external morphology but complex jaw apparatus. The structure of jaws has been traditionally used in the systematics of the family in the generic diagnoses. Polina is utilizing the methods of microCT to study the jaws of lumbrinerids in 3D.
Jaws of Scoletoma fragilis from the White Sea scanned using microCT showing ventral solid mandibles, forceps-like maxillae I and denticulate maxillae II and II, carriers of maxillae are omitted (Photo: P. Borisova)
Polina is also going to sequence several genetic markers to reconstruct the first molecular phylogeny of the family. This will allow testing the current hypothesis on the intergeneric relationships within Lumbrineridae and will aid in tracing the evolution of jaws within the family.
Molluscs come in a variety of shapes and sizes, but some of the least known are perhaps the Aplacophora, or shell-less molluscs. Instead of a shell, these worm-shaped molluscs have a cuticle covered in calcareous spicules, or sclerites, that give them a beautiful, glistening appearance!
The very first species of aplacophoran mollusc, Chaetoderma nitidulum, was collected from the Swedish west coast and described by the Swedish taxonomist Sven Lovén in 1844. At the time, it was not even known what animal group the new, strange animal belonged to. It had spicules– could it be related to the spiny sea urchins? It had a worm-like body– could it be related to other worm-shaped animals? It would be almost 50 years before it was conclusively recognized as part of Mollusca. Since then, many more species have been discovered, and today close to 500 species of aplacophoran molluscs have been described.
A specimen of Chaetoderma nitidulum from the Norwegian West Coast Photo: N. Mikkelsen
Chaetoderma nitidulum is known today as one of the common aplacophoran molluscs in the East Atlantic, with a distribution from the Svalbard archipelago in the north, to the British Isles in the south. However, taxonomist have been debating the identity of Chaetoderma nitidulum since shortly after it was described. Some researchers have suggested that it could in fact consist of up to six different species. Other researchers have synonymized it with other species, or suggested that it is not a separate species, but only part of a larger species which has a distribution that spans the entire North Atlantic.
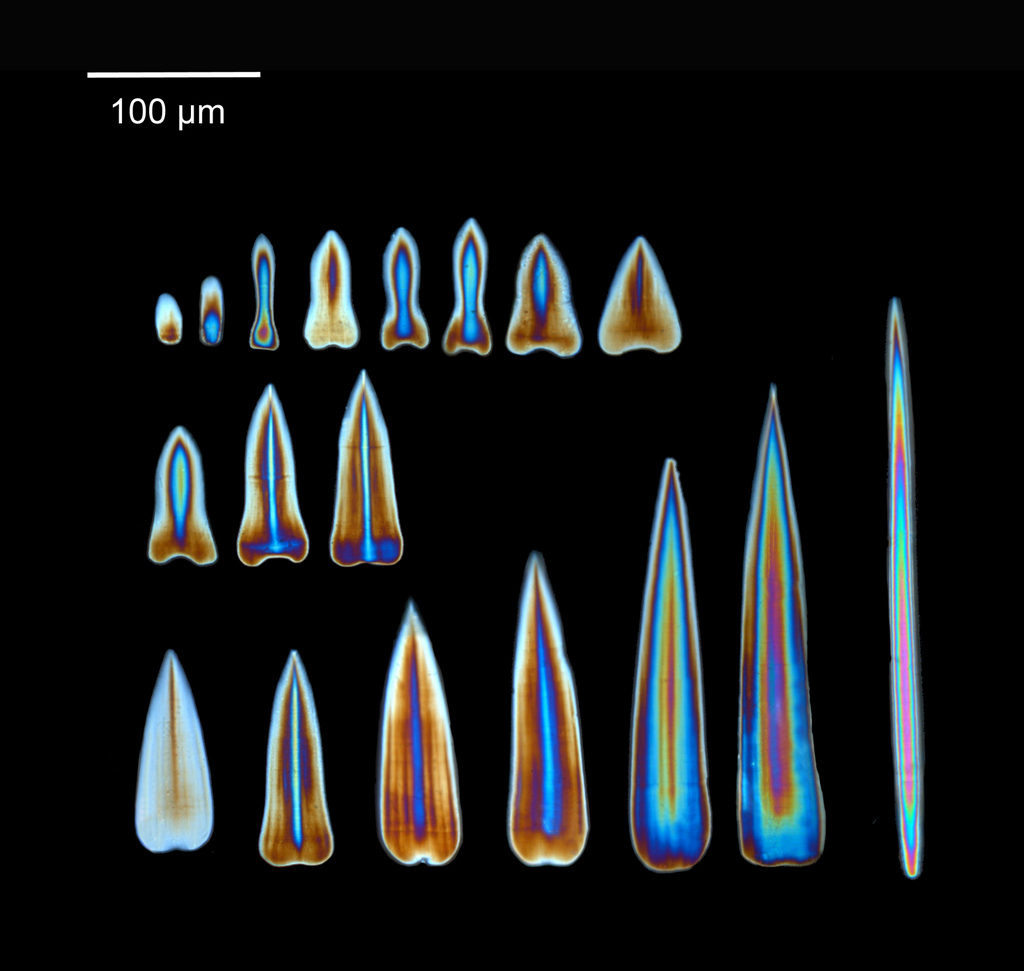
The shape, size and the patterns on the calcareous sclerites covering the body of the aplacophoran molluscs is unique to each species, making it one of the most important characters we have to distinguish between different species.
Calcareous clerites from Chaetoderma nitidulum. Photo: N. Mikkelsen
Looking at the sclerites through the microscope equipped with a cross-polarizing filter gives us a shiny, colorful view of the sclerites. The light shines with different colors depending on the thickness of the sclerites, helping us get a good view of the structure of the sclerites.
Sclerites from Chaetoderma nitidulum viewed under cross-polarized light. Photo: N. Mikkelsen
We have recently investigated specimens of Chaetoderma nitidulum from different localities from the entire distribution range of the species. Our investigations have revealed a lot of variation between the specimens, both in the calcareous sclerites and in DNA sequences, separating the specimens into at least two different groups. Could it be that Chaetoderma nitidulum actually represents more than one species?