This year started with an exciting adventure, the kind that most marine biologists dream about since we were kids: an Antarctic expedition.
As part of the Manet Team’s ongoing projects exploring the diversity of polar jellies and polyps, we joined the Challenge-2 Team from University of Barcelona led by Elisenda Ballesté and Conxita Ávila, a fantastic group of highly motivated researchers from all career stages accompanied by expert divers taking good care of the challenging underwater operations. The goal: evaluate the anthropogenic impacts that can affect wildlife in this pristine environment. For Joan, this was the second time in the white continent, but the prospects of diving in its glacial waters and witness their underwater wonders was beyond exciting. For Luis, this was his first time, and not only he found tones of interesting species and enjoyed birdwatching rare Antarctic species, but also had to “fight for his life” against a very brave, young sea lion, quite curious about the intertidal work he was doing.

We step on board the Spanish research vessel BIO Hesperides in Ushuaia (Argentina) at the beginning of January, crossed the Drake Passage in just a few days, and spent most of the 6-weeks cruise around the magnificent west side of the Antarctic Peninsula and Orkney Islands, before disembarking in Punta Arenas (Chile) by mid-February. The breathtaking Antarctic scenery was complemented by the diverse wildlife that calls this region home. We were fortunate to observe a multitude of seabirds like majestic albatrosses, playful petrels, not-so-playful Antarctic skuas, and the enigmatic snowy sheathbill. We also encountered several species of penguins —including the iconic Gentoo and Adélie penguins— and marine mammals —including a myriad of humpback whales, the spine-chilling leopard seals, massive sea elephants, and curious crab eater seals—which were a delight to watch as they waddled across the ice and slid into the water. Their graceful and sometimes funny movements were a constant reminder of the rich biodiversity thriving in these remote waters, while adding a touch of magic to our scientific endeavors.

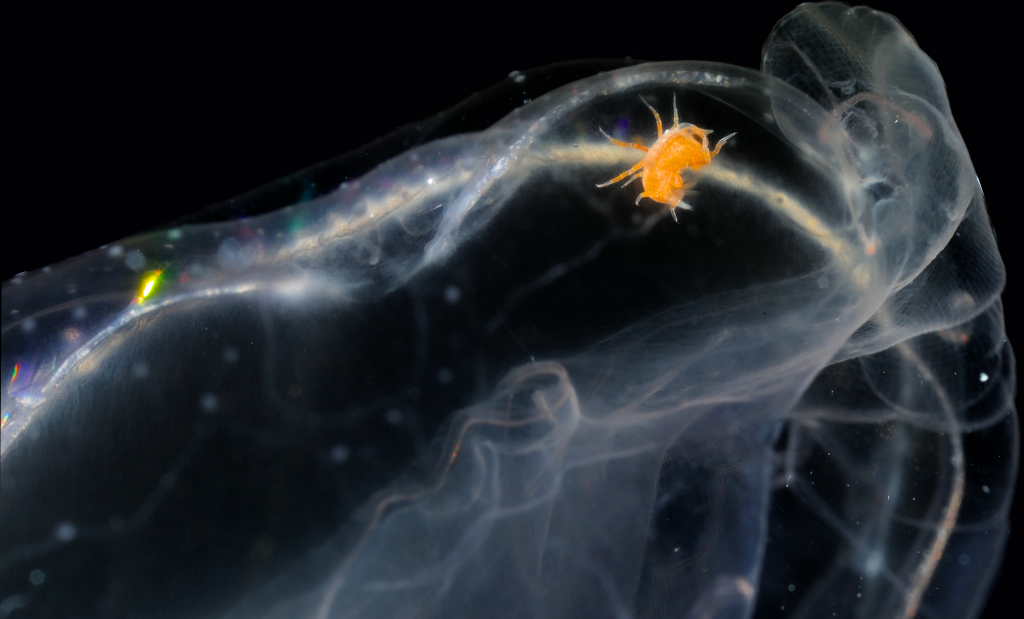
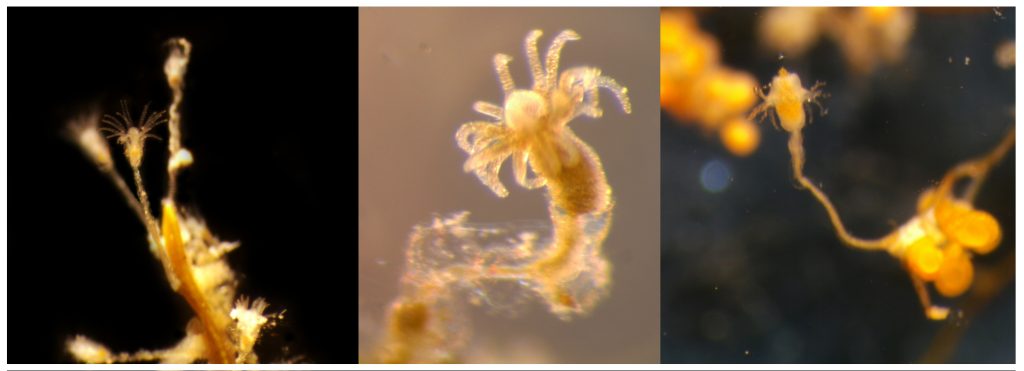
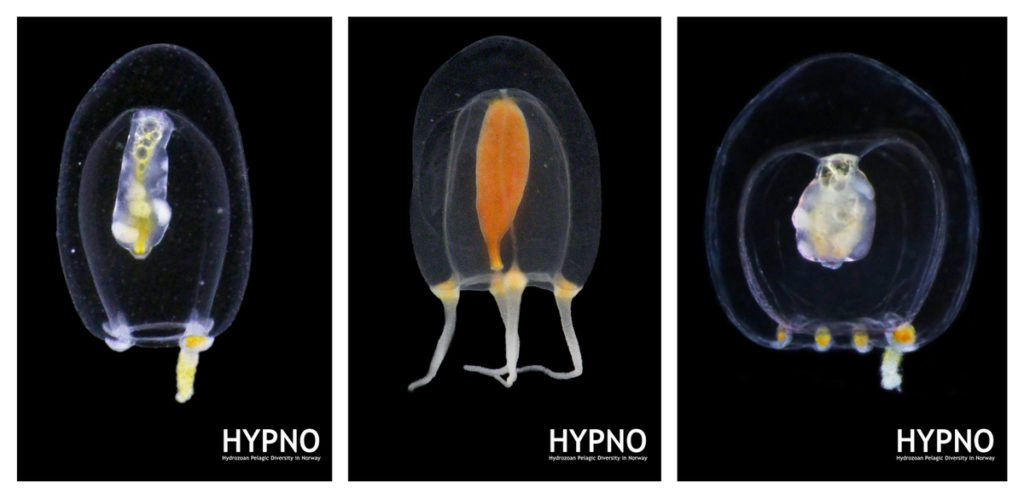
However, the creatures that truly captured our excitement were, of course, the jellyfish and polyps we encountered. Prior to throwing our nets (or ourselves) into the cold unknown, we had to carefully plan our sampling protocols and set realistic, yet ambitious goals. Obtaining high-quality samples in one of the most isolated and extreme environments on Earth is certainly a challenge, but when done adequately and in a safe way, it can also be lot of fun. We targeted both planktonic stages (hydromedusae, siphonophores, ctenophores and tunicates) we collected using a brand new WP3 net, and benthic ones (hydroids) we collected while diving in icy waters around the freezing point.

We collected over 50 different species of jellyfish and polyps, some of which had never been documented in this region before. Each specimen we retrieved was meticulously photographed and cataloged. These fascinating organisms, often overlooked in the grand tapestry of marine life, play crucial roles in the ecosystem. But how are we going to evaluate potential impacts in the ecosystems if we don’t even have a good knowledge of the true diversity that thrives in these extreme environments? That’s precisely both our mission and the reason why we had the chance to join this fantastic opportunity. Our small team of two focused on studying the diversity and distribution of Hydrozoa, as part of our project POLE2POLE (MSCA, Horizon2020), which investigates the phenomenon of bipolarity in the group. The data we gathered will not only help us comprehend the ecological roles these organisms play, but also shed light on how they may be impacted by anthropogenic factors such as pollution and climate change in the (very) near future.

In addition to exciting material for our research projects, the expedition provided us with invaluable opportunities to engage with our comrades at Challenge-2 and the scientific community on board. Collaborating with researchers from various institutions, we discussed the implications of our research and explored ways to mitigate the impacts of human activities on this fragile ecosystem. As we concluded our expedition, we left with a profound appreciation for the Antarctic’s beauty and the complexity of its ecosystems, as well as some seed ideas for potential future research projects in the area. The experiences shared on board the BIO Hesperides, from witnessing the majestic wildlife to uncovering the hidden diversity of jellyfish and polyps, reinforced our commitment to studying and preserving this remarkable region. Through projects like Challenge-2 and POLE2POLE, we hope to contribute to a greater understanding of the Antarctic diversity, and advocate for its conservation for generations to come.
Joan and Luis, Manet Team