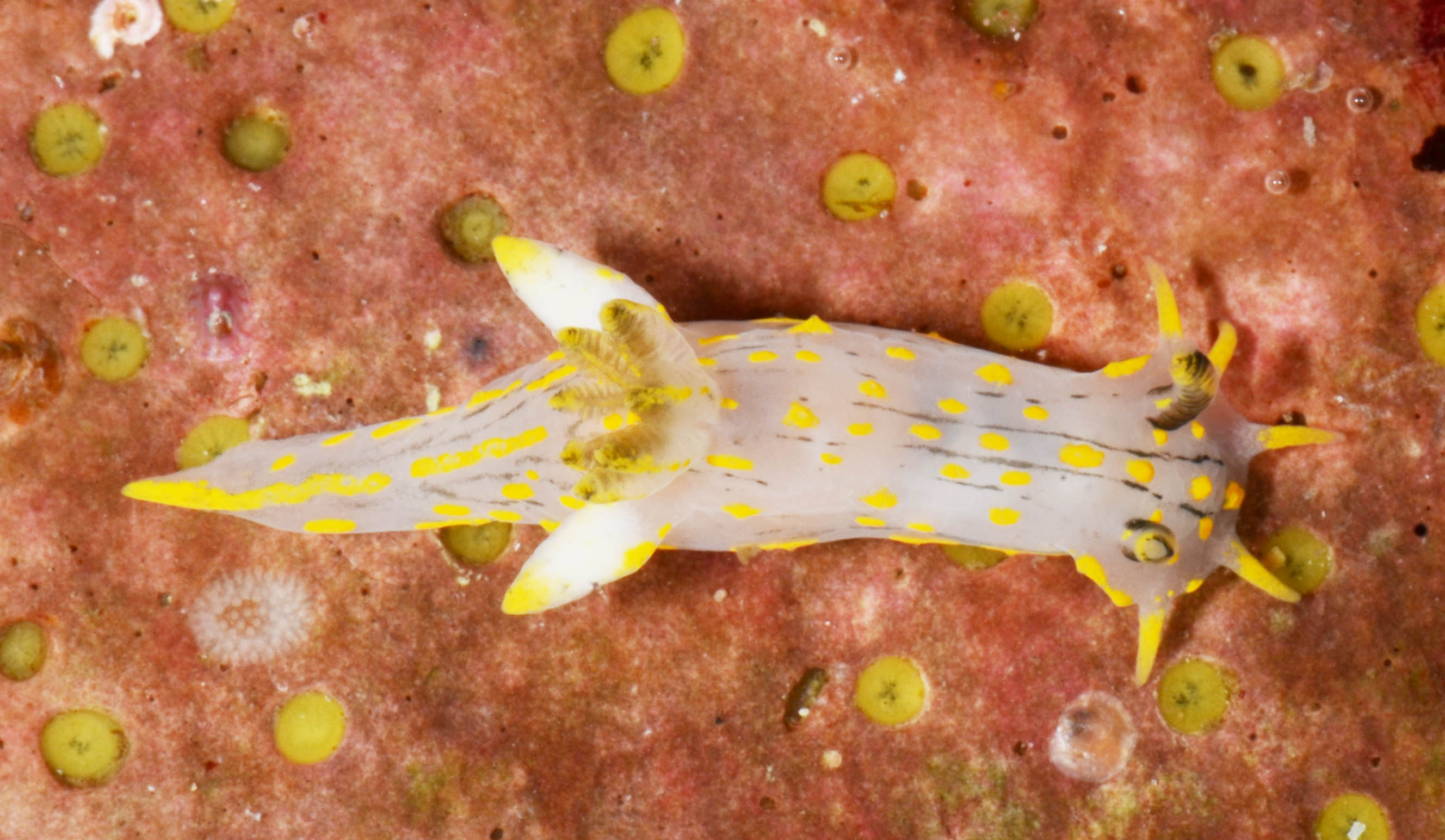
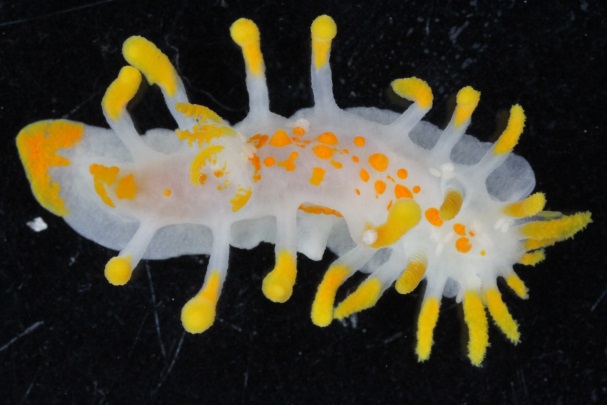
Polycera quadrilineata (Norway) Photo: M. Malaquias
Phillidia ocellata (Mozambique) Photo: M. Malaquias
Chromodoris cf. quadricolor. Vamizi Island
Elysia subornata (Key Largo)
Chromodoris africana (Zavora, Inhambane). This species is part of a complex in need of revision where other “species” imaged here are also part of (e.g. Chromodoris hamiltoni, Hypselodoris regina, Chromodoris elisabethina)
Untangling the diversity and evolution of Sea Hares
Aplysia parvula; Føllingen, Norway; Photo by Nils Aukan
Sampling and freezing at Askøy
Dr Carlo M. Cunha from the Metropolitan University of Santos in Brazil (Universidade Metropolitana de Santos), a world expert in the diversity and systematics of Anaspidea heterobranch gastropods, visited the Natural History Museum of Bergen for a month during January/February 2017 to study our scientific collection of these molluscs. The visit was funded by the University of Bergen´s Strategic Programme for International Research and Education (SPIRE).
The Museum holds a large amount of material from the Scandinavian region, but also from the Mediterranean, Macaronesia islands, Caribbean, and western Indian Ocean.
These marine molluscs commonly known by sea hares comprise around 90 currently known species and have long been of major interest to biologists because of their large and easily accessible nervous system, which form the basis of numerous neurophysiological works.
Preserved specimen of Aplysia punctata from Norway
Dissected specimen of Aplysia punctata from Norway
However, the taxonomy of these molluscs and their evolution are still poorly understood. Dr Cunha is using a combination of molecular and morphological tools to learn more about the worldwide diversity of anaspideans and their phylogenetic relationships.
Dr Cunha visit to Bergen has already resulted in the revision and update of the taxonomy of our Anaspidea collection. The Norwegian species of anaspids were revised and redescribed in detail using electron microscopy and DNA barcoding performed in collaboration with Louise Lindblom (University Museum / Biodiversity Labs).
SEM-image of jaws of Phyllaplysia sp from Florida, USA
Additionally several other species from around the world were studied and will be integrated in ongoing taxonomic revisions. Keep tuned!
-Manuel
We’ve also had Lloyd visiting recently, you’ll find a post about that on the Marine Invertebrates of Western Africa blog: click here
A whale recently had to be put down by wildlife management after it had repeatedly beached itself on the island of Sotra outside of Bergen. It was found to be a Cuvier’s beaked whale (Ziphius cavirostris), a species with apparently no official previous records from Norway. The University Museum of Bergen therefore wished to include the whale skeleton in its collections (and future exhibitions, once the remodelling completes).
Arriving at Espegrend
The whale was transported to the Marine Biological Station of Espegrend, and a team of five people from the museum set to work collecting measurements of the whale, taking tissue samples for DNA-barcoding though the NorBOL-project, collecting ectoparasites, and doing photo-documentation.
Collecting measurements
We then began removing the blubber and muscle tissue off the whale so that the bones can be further treated (they contain a lot of oil which needs to be taken care of once the soft tissue has been removed), before the skeleton can be mounted for display.
Starting the work of removing blubber and muscles
Little did we know that what had so far been a local news matter would soon go viral…
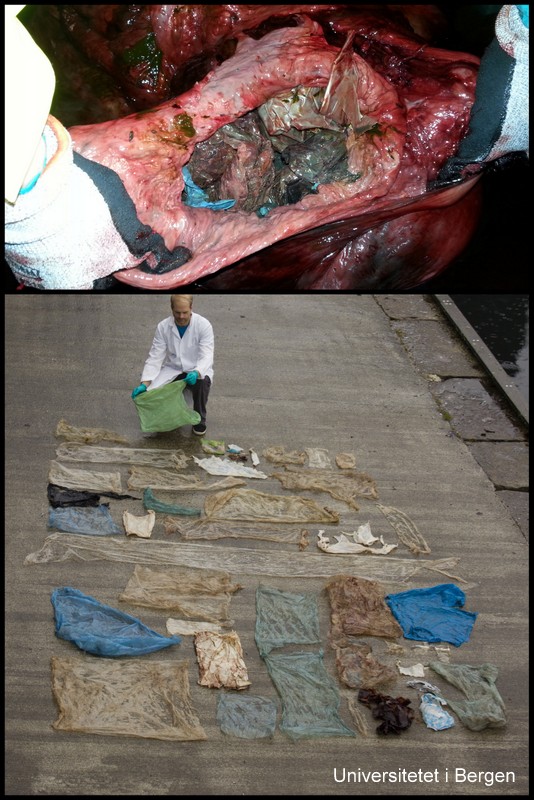
Sadly, it became clear during the autopsy that the whale had been ingesting massive amounts of plastic – as much as 30 plastic bags, and many smaller pieces of plastic. The whale was emaciated, and we believe that the plastic had gathered in such an amount in its stomach that it had created a plug, stopping the digestive process.
The plastic in and from the whale stomach (photos: T. Lislevand, H.Glenner/C.Noever)
The images of all the plastic spread out on the ground became a potent reminder of the tragedies that marine pollution is creating, and has sparked a renewed debate on how we can limit the amount of micro- and macro-plastic that end up in nature.
The news of the whale’s stomach content became international news
What should the Cuvier’s beaked whale have been eating?
Occurring as solitary animals or in small pods, and preferring the deeper open waters, the Cuvier’s beaked whale is not an easy animal to study. We do know that the species have a more or less cosmopolitan distribution, and that it holds the world record for longest and deepest dive for any mammal: one was recorded diving down to 3000 meters.
What data we do have on the species diet comes from beached individuals, and suggests that the species may be a fairly omnivorous predator. From the limited number of Cuvier’s beaked whales that have been examined for stomach content, there are regional differences in the diet, but it seems to consist mainly of cephalopods (squid and octopuses), deep sea fish, and medium sized crustaceans (Santos og andre 2001).
Above are the suckers on the arm of a giant squid, Architeuthis. Below are scars on the skin of a sperm whale. Photo: E.Willassen
The cephalopods appear to be the dominant food source, but this interpretation may be influenced by the longevity of the hard parts of a cephalopod in the stomach.
The tough beaks of a cephalopod consist of chitin, and is used for tearing prey to pieces. Chitin is also found in the suckers of many cephalopods. The beaks can be used to identify the cephalod groups based on their size and shapes. Animals such as jellyfish would be much harder to document as part of the diet, as they would be digested much more rapidly and completely.
We don’t know how well resolved the information produced by the animal’s echo-location is, but it is conceivable that the plastic reflects signals in a way similar to the natural food of the whale, and is therefore “caught” and eaten.
Cephalopod beak, drawing by J.H. Emerton (from Wikimedia commons)
We did find some cephalopod beaks in between the plastic in the whale stomach – so far we have not had the time to attempt to identify these, but we will.
Amongst the plastic there are some cephalopod beaks (dark brown) and a bivalve shell (top left). Photo: C. Noever
The University Museum have extensive cephalopod collections, and long traditions for working with this group – from Dr. Jakob Johan Adolf Appellöf who began working here in 1890, to the material collected in the MAR-ECO project.
MAR-ECO workshop on cephalopoda
From the work of Santos et al 2001 we know that the following species are in the diet of European Curvier’s beaked whales, and are probably amongst the things our whale should have been eating:
Tewuthowenia megalops. Photo: Richard E. Young during MAR-ECO-cruise 2004.
Teuthowenia megalops is an odd squid that floats around in the open water with a propulsion system based on ammoniumchloride that the animal produces by digesting protein. The name “megalops” hints to the huge eyes, which also contain three light producing organs (chromatophores). The species seems to be common in deep water in the north Atlantic (Vecchione et al. 2008). For more information, see Wikipedia.
Mastigoteuthis agassizi
Mastigoteuthis agassizii was originally registered in whale stomachs as Mastigoteuthis schmidti, but from the work on the MAR-ECO project, three species of Mastigoteuthis were considered to all be M. agassizii. Some ambiguity remains about the species of this genus of oceanic squid with a broad distribution in the world’s oceans in depths ranging from 500 to 1000 meters. They have diurnal migration, and may be found hunting closer to the surface at night.
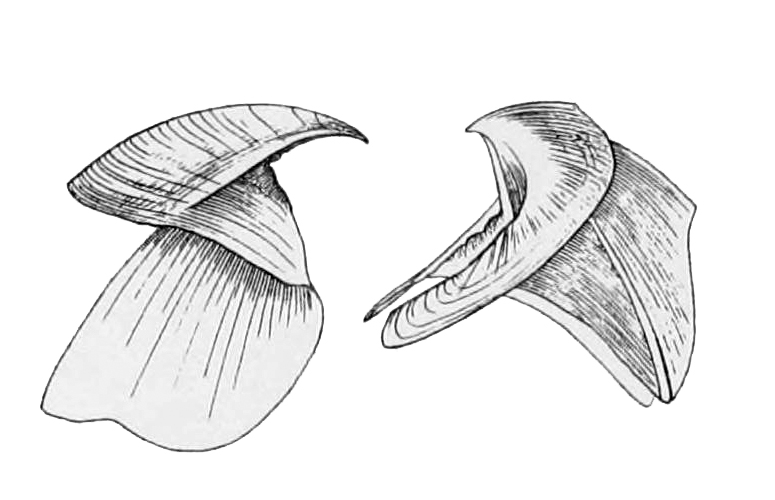
Taonius pavo seen ventrally (above) and dorsally. Illustration from Wikipedia.
Taonius pavo
This little squid is not very well known. It has been recorded from the Atlantic Ocean, but it may have a broader distribution. In this link you will find a video from the Bahamas at 850 m depth where the animal releases bio- luminescent “ink” to confuse a predator and escape.
Histioteuthis bonelli Photographed by Richard E.Young during the Mar-Eco-cruises in 2004
Histioteuthis bonelli, drawing by Ernst Haeckel.
Histioteuthis bonnellii has several names in English, one of which is “umbrella squid”. The name is due to the skirt-like membrane between the arms – when it splays its arms it resembles an umbrella. We don’t know much about the biology of H. bonellii, except that it has several close relatives in the world oceans, and that what has hitherto been considered one species (H. bonellii) may well turn out to be several species.
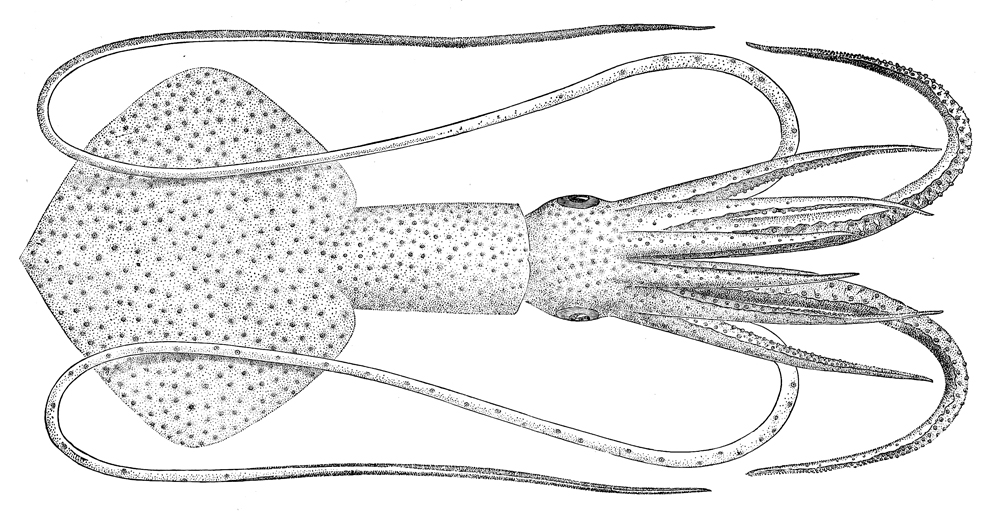
Todarodes sagittatus
Todarodes sagittatus, the European flying squid, is one of the ten-armed cephalopods that may irregularly occur in schools along the Norwegian coast. T. sagittatus is subject to fisheries.
Vampyroteuthis infernalis
Vampyroteuthis infernalis – the vampire squid is a deep-sea squid with eight arms and a skirt-like mantle between its arms. It also has moveable wings on its body that it can use to manoeuvre with. The name “vampire squid” is not quite true – this is no blood sucker, but it traps organic material from the water masses using long, sticky threads. If threatened, it can invert the “skirt” over its head, resembling a hedgehog. It also has light producing organs towards the back of the body, and can create clouds of bioluminescence. Even with all these defences, it may end up in the stomach of a Cuvier’s beaked whale.
Pelagic crustaceans and deep sea fish are also amongst the recorded prey from Cuvier’s beaked whales. Amongst these we find the fairly large and shrimplike Gnathophausia, found within the order Lophogastrida, which has been studied extensively at the University of Bergen. We also found a bivalve shell in the stomach of our whale, which as far as we are aware of has not been recorded as part of their diet previously.
Plastic or food?
It may seem strange that the whale should ingest large amounts of plastic – why would it do that? If the whale primarily finds its pray by echolocation in the pitch black of the deep sea, it may well be that it is unable to differentiate between the reflected signal from a sheet of plastic, and that from one of its usual prey animals.
Unlike the sperm whales that hunt cephalopods in a similar way, the beaked does not have teeth to grab its pray. Instead they use a suction to ingest the food. Perhaps it is this feeding mode that becomes very unfortunate for the whales in a natural environment with an incredible amount of human garbage.
Hyalinoecia tubicola from the North Sea (by K. Kongshavn).
Quill worms belong to the annelid family Onuphidae and are called like that because of their unique tubes. The tubes are secreted by their inhabitants and are very light and rigid, resembling a quill, the basal part of a bird’s feather used for writing. Quill worms are epibenthic creatures capable of crawling on the surface of the sea floor carrying their tubes along. Their anterior feet are modified, strengthened and enlarged, bearing thick and stout bristles. These anterior feet are used for locomotion.
Quill worms are widely distributed in the ocean inhabiting mostly slope depths down to 2000 m. Being large in body size (up to 10-20 cm long), they can be quite abundant in some areas. Meyer et al. (2016) reported Hyalinoecia artifex reaching up to 70 ind./m2 in the Baltimore Canyon at 400 m water depth. Another quill worm, H. tubicola, which is very common in Norwegian waters, reached up to 272 ind./m2 at 365 m offshore of Chesapeake Bay (Wigley & Emery 1967).
Quill worms are believed to be motile scavengers. Baited monster camera experiments performed at 2000 m deep site in Baja California demonstrated that Hyalinoecia worms can accumulate in hundreds of specimens five hours after the bait (rotten fish) has been deployed (Dayton & Hessler 1972). Myer et al. (2016) analyzed the stable isotope content in Hyalinoecia artifex tissues confirming its secondary consumer status. Their results supported earlier observations on the gut content of the same species by Gaston (1987) showing the presence of the remains of various benthic invertebrates.
Video 1. Quill worm Hyalinoecia tubicola moving inside its tube (by K. Kongshavn).
Video 2. Quill worm Hyalinoecia tubicola protruding from the tube opening. Three antennae and a pair of palps are seen on the head. The first two pairs of feet are enlarged and strengthened (by K. Kongshavn).
Dayton, P.K., Hessler, R.R., 1972. Role of biological disturbance in maintaining diversity in the deep sea. Deep-Sea Research 19: 199–208.
Meyer, K.S., Wagner, J.K.S., Ball, B., Turner, P.J., Young, C.M., Van Dover, C.L. 2016. Hyalinoecia artifex: Field notes on a charismatic and abundant epifaunal polychaete on the US Atlantic continental margin. Invertebrate Biology 135: 211–224. doi:10.1111/ivb.12132
Gaston, G.R. 1987. Benthic polychaeta of the Middle Atlantic Bight: feeding and distribution. Marine Ecology Progress Series 36: 251–262.
Wigley, R.L., Emery, K.O. 1967. Benthic animals, particularly Hyalinoecia (Annelida) and Ophiomusium (Echinodermata), in sea-bottom photographs from the continental slope. In: Deep-Sea Photography. Hersey JB, ed., pp. 235–250. John Hopkins Press, Baltimore.
Today we present two more of Arne Nygrens gorgeous photos, that he made during our week in the field in Sletvik (central Norway). The subjects in both of these are polychaetes from the family Phyllodocidae, the paddleworms.
First up is a stunning Phyllodoce citrina collected from shell sand at about 60 m depth. The animal is approximately 6 cm long.
Phyllodoce citrina, Photo by Arne Nygren CC-BY-SA
The next one, Paranaitis sp. n. is actually a new species for science, which came as a pleasant surprise. This is a fairly well-studied group, and the locality Galgenes is one that has been sampled regularly – yet there it was! It is rather unusual to find species where one can so immediately recognize that they are something new; usually we need many specimens, and a combination of detailed studies of morphology and genetic work – but this one is possible to distinguish straight from morphology, as it was lacking eyes. The specimen is about 1.5 cm long.
Last year we had a calendar post about the Heart of the Museum – our type collections.
To recap, a species’ type is “…the objective standard of reference for the application of zoological names. When a new species or subspecies is described, the specimen(s) on which the author based his/her description become the type(s) (Article 72.1). In this way names are linked to type specimens, which can be referred to later if there is doubt over the interpretation of that name.
Consequently types are sometimes referred to as “onomatophores” which means name bearers.”
The location – sampling site – from which the type specimen is described is known as the type locality.
Michael Sars (image from Wikimedia)
As you have probably noticed, polychaetes (bristle worms) are a focus group in our lab, and several species have type localities close by.
The biologist and theologian Michael Sars (1805-1869) lived in the Bergen region for many years. He was a prolific taxonomist, naming 277 species of marine taxa according to the World Register of Marine Species (WoRMS).
Consequently there are quite a few species that have their type locality within easy daytrip-distance by ship for us.
On the hunt with R/V “Hans Brattstrøm”
One such locality is Glesvær, where Michael Sars described several new species in his work of 1835: Beskrivelser og Iagttagelser over nogle mærkelige eller nye i Havet ved den Bergenske Kyst levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes og Molluskernes Classer* (“Descriptions and Observations of some strange or new animals found off the coast of Bergen, belonging to the Classes …”).
The polychaete Amphicteis gunneri (Ampharetidae) is one of these species. It was first described by Michael Sars as Amphitrite gunneri (the species name is an homage to Johan Ernst Gunnerus (1718-1773) who was an active scientist within botany and zoology, as well as the bishop in Trondheim, and one of the founders of Det Kongelige Norske Videnskapers Selskap) in the publication above. Here are his original illustrations of the species:
Amphicteis gunneri by M. Sars (1835)
We have previously submitted several specimens of Amphicteis gunneri for DNA-barcoding through the NorBOL-project – and found that specimens that according to the keys in the literature should all come out nicely as A. gunneri in fact end up in several barcode-based groupings (BINs), meaning that they genetically different from each other. Then we need to unravel which one is the true A. gunneri, and decide what to do with the others. In such cases, material from type localities is invaluable. By sending in specimens identified by resident taxonomists as A. gunneri from the type locality, we hope to figure out which BIN represent A. gunneri, and which represent potentially new species.
We were also able to photograph live specimens showing the nice coloration of this worm. Fixed specimens lose this colour and become uniformly yellow/white (no dots).
Amphicteis gunneri collected at type locality. Photo: K.Kongshavn
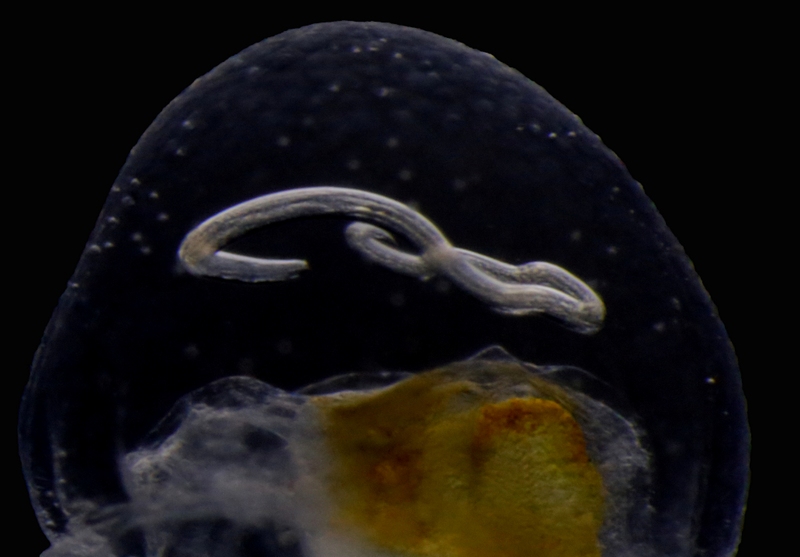
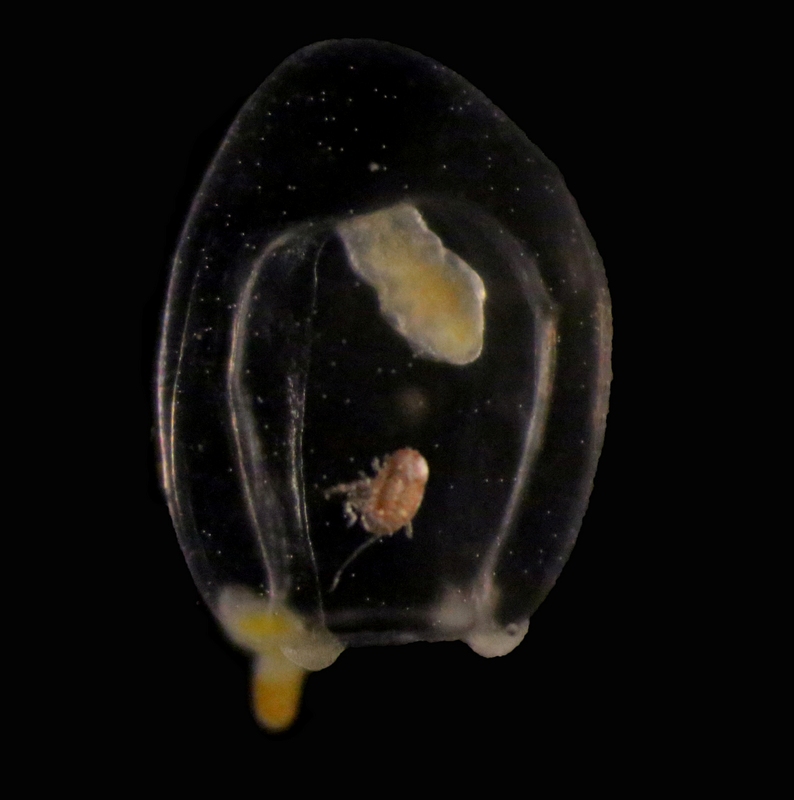
Travelling alone through the water column may be a dangerous business: reaching the final destination is not always guaranteed, the risk of being eaten is high, and even finding food may prove a difficult task… which is why several animals choose to travel comfortably on or inside jellyfish and siphonophores!
Jellyfish are commonly involved in relationships of parasitism and phoresis (i. e., when one organism is mechanically transported by another without any further physiological dependence), and many examples have been observed of these interactions around the world. For instance, it’s not unusual to find hyperiid amphipods and sea-spiders –as well as lobster and crab larvae – piggybacking on the surface of large and tiny jellyfish, and while it’s still not clear whether or not all these passengers feed on their means of transportation, real parasitism and jelly-feeding has been confirmed for at least some of them. Jellyfish may also transport parasitic worms to their final hosts (like the nematode you see in the pictures), acting as carriers of parasites towards fish and mammals, and sometimes, eventually reaching humans as well!
Euphysa aurata medusa with parasitic nematode larva. Korsfjord, February 2016. Credit: Aino Hosia.
A close-up of 2 showing the parasite embedded in the mesoglea (jelly) of the host. Credit: Aino Hosia.
Euphysa aurata medusa with crustacean ectosymbiont. Raunefjord, December 2016. Credit: Luis Martell
These two hydromedusae of Euphysa aurata were collected this year in the fjords south of Bergen, and are only an example of jellyfish harboring other animals in this area. The species is a common and widespread jellyfish around here, but its role in the transmission of parasites and transportation of small crustaceans has never been explored. It might well be that, together with its gelatinous relatives, E. aurata will prove to be involved in many more biological interactions than we previously thought!
Today, I thought I’d introduce to you to a cool group of animals that is ubiquitous in the oceans (including the Norwegian seas), but unfamiliar to most people. Siphonophores (“kolonimaneter” in Norwegian) belong to cnidarians, a group that includes corals, anemones, hydroids and jellyfish, and is characterized by the presence of stinging cells used in prey capture. All siphonophores are predatory, and use their stinging tentacles to catch small crustaceans or, in the case of some species, even small fish.
The most (or only) familiar siphonophore for the majority of people is probably the highly venomous Portuguese Man O’War (Physalia physalis), which can be spotted floating on the surface of the ocean or stranded on beaches. However, it is not really representative of the group as a whole, as most siphonophores live in the water column of the open ocean rather than its surface. There are around 200 described species of siphonophores.
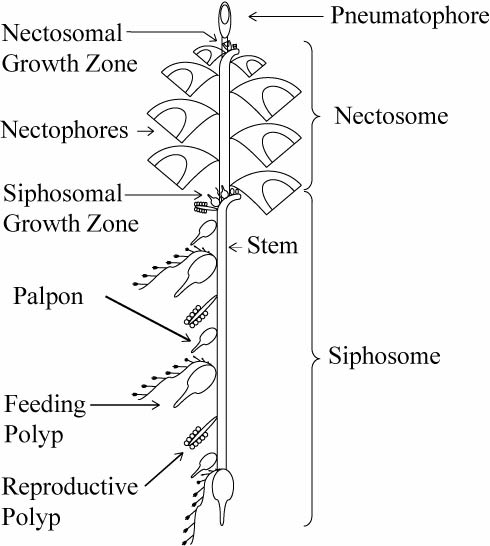
The most fascinating feature of siphonophores is their peculiar body plan. While siphonophores may appear to be a single animal, they are in fact a colony of physiologically connected and genetically identical but morphologically diverse individuals called zooids that have specialized to carry out different tasks for the colony. Siphonophores belong to the class Hydrozoa (“polyppdyr” in Norwegian), which covers two basic body plans: the polyp/hydroid and the medusa.
Schematic of a physonect siphonophore. From http://www.siphonophores.org (CC-by-nc-sa)
The various zooids comprising a siphonophore colony can also be divided into these main groups. For example, the zooids used for swimming, called nectophores, are medusoid, while the feeding zooids, or gastrozooids, are polyp-like. The siphonophore colony can also include specialized defensive, protective and reproductive zooids. All the zooids forming a colony arise by budding from a single fertilized egg. The different zooids are specialized to the degree that they cannot function as individual animals any more, and are only able to perform their specific tasks as parts of the siphonophore colony.
Anterior nectophore, posterior nectophore and eudoxid of the calycophoran siphonophore Dimophyes arctica – a common species in Norwegian waters. Photos by Aino Hosia (cc-by-sa)
The zooids, for example the swimming nectophores, vary in appearance between species, and can be used for species identification. In addition, the various types of zooids in the colony are arranged in a strict species specific pattern, providing the intact colonies of each species with their particular appearance. While the individual zooids are generally small, millimeters to centimeters in size, some siphonophore species, like Praya dubia, may have colonies that reach 40 m in length! Siphonophore colonies generally have a zone of one or more (up to several dozen) swimming nectophores at the front, used to pull the colony through water. Behind this nectosome is the siphosome, which contains the feeding, reproductive and other zooids in a repeating pattern, each iteration of which is called a cormidium. In some species (suborder Calycophorae), these cormidia are released as small free-living reproductive colonies called eudoxids. Unfortunately, siphonophore colonies are extremely fragile and tend to fall apart during standard plankton sampling with nets, leaving behind a bewildering array of small bits and pieces – part of the reason they are relatively poorly known to most people.
Colony of physonect siphonophore Physophora hydrostatica, aka hula skirt siphonophore. Photo by Aino Hosia (cc-by-sa)
Intact siphonophore colonies are beautiful, but often utterly alien in appearance. It is interesting to consider where to draw the line between an individual and a colony. While we as individuals have specialized organs to carry out our various bodily functions, siphonophore colonies are made up of specialized interdependent individuals or zooids similarly carrying out their specific tasks.
As part of project HYPNO we are charting the diversity of pelagic hydrozoans, including siphonophores, in Norway. There are ~15 species observed in Norwegian waters, and some, particularly Dimophyes arctica, Lensia conoidea and Nanomia sp. are extremely common components of marine plankton. However, siphonophores are primarily noticed when they become a nuisance: For example, mass occurrences of Muggiaea atlantica and Apolemia uvaria have in the past killed large numbers of farmed fish in Norway, with resulting losses to aquaculture companies.
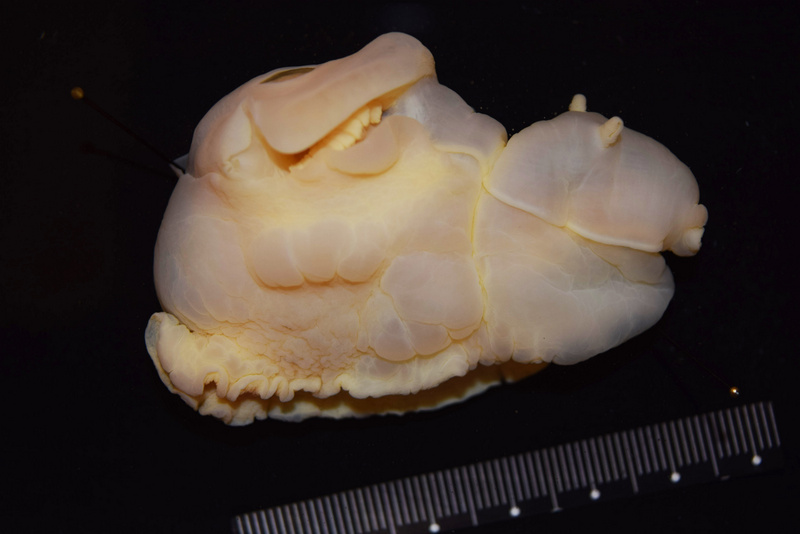
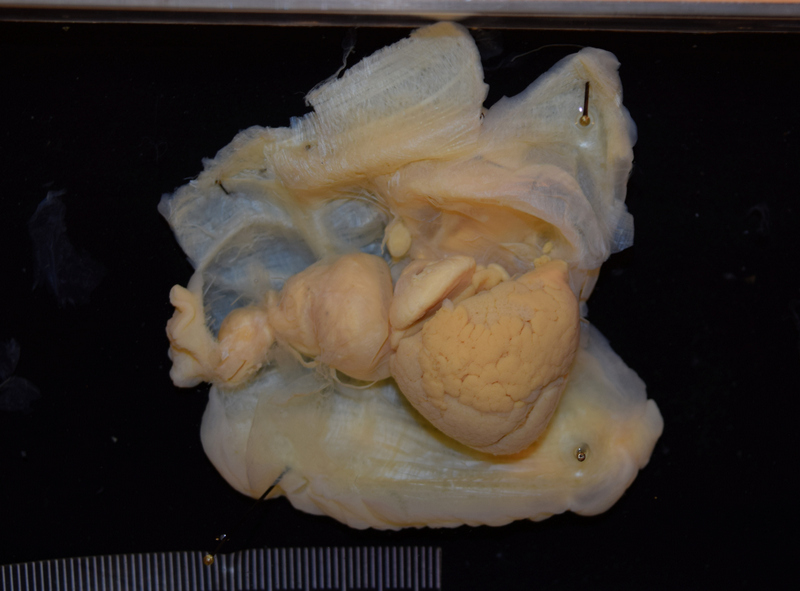
During my stay at the University Museum of Bergen I have been working on the diversity and systematics of Antarctic philine snails. Most of the samples were collected during different cruises on board of the RV Polarstern in the Eastern Weddell Sea, Bouvet Island, and South Shetland Islands (West Antarctica). I photographed all specimens and then clipped them for the DNA analysis (see pictures).
Philine specimens
I was able to work at the DNA lab with excellent resources for DNA extraction, amplification, purification, and sequencing.
I am indebted to Louise Lindblom who helped me at the beginning of my crusade there. After a first barcoding of all the material we identified six clades, from which we selected a maximum of three specimens to further sequence the ribosomal genes 16S and 28S and the nuclear gene codifying for the Histone 3.
The first phylogenetic tree with all partitions resulted in the finding of novel clades that now deserve further investigation.
Prof. Manuel António E. Malaquias and his PhD Student Trond Oskars helped me dissecting the material for anatomical analyses. Important taxonomical characters were those related to the male reproductive system, the digestive tract as well, and the shell. After the dissections and drawings of the main parts I prepared the hard structures such as the radula, the shell, and the gizzard plates for Scanning Electron Microscopy (SEM) as well as some soft structures after critical point drying. I could photograph all these material at the same facilities of the museum being helped by Irene and Katrine. After the two months of work, I ended up having huge amount of anatomical and molecular data that deserves further processing. See a picture of the radula and a gizzard plate:
Gizzard plate
Radula
Moreover, I was able to join the student diving club and make several dives to get to know the local flora and fauna. I could even collect some other heterobranch slugs for the barcoding project of the museum. See a couple of pictures of the nudibranch Limacia clavigera and Onchidoris muricata.
Onchidoris muricata
Limacia clavigera
Overall, Bergen is a nice city to visit surrounded by nice mountains, good (but not cheap) beers, beautiful fjords, and nice people. I hope I can come back with a postdoctoral position to further enjoy the country and meet more Viking descendants.
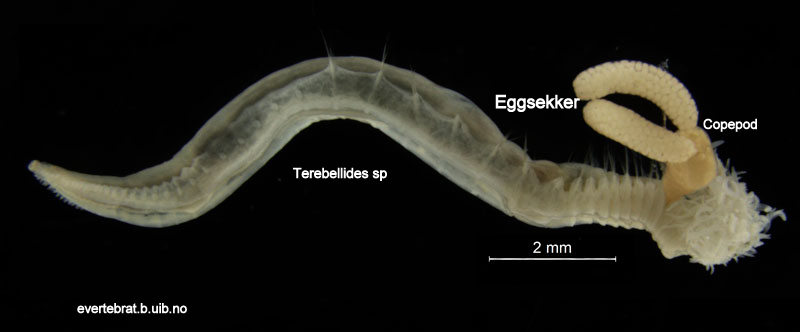
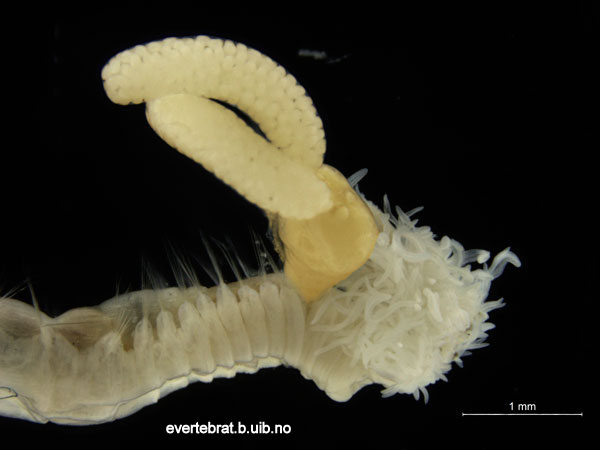
Today is #WormWednesday on Twitter, and we figured that it was a good day to introduce you to this rather unlucky fellow and his sidekick…
The orange part (coloured in Photoshop) is the parasite. The two long sacks are filled with eggs.
They were collected during our field work in Sletvik in October. The worm is a polychaete in the genus Terebellides, whilst the parasite is a Copepod. This species rich group of small crustaceans have many modes of life, but parasitism is a common one, with about half of the ~13 000 species being parasites.